
Titelbild: 3D Modell des Schädels von Captorhinus aguti, einem urtümlichen Reptil aus dem Perm der USA. Finden sich in ihm mögliche Vorstufen für Schläfenöffnungen? Aus Abel et al. (2022).
In den ersten beiden Teilen (1 & 2) haben wir einen Rundumschlag zur Frage der Schläfenöffnungen gemacht. Wir haben uns sowohl angesehen, wie verschiedene WissenschaftlerInnen versucht haben Ordnung in die Vielfalt der Schläfenöffnungen zu bringen, als auch welche biologischen Gründe dieser Vielfalt zugrunde liegen könnten. Als wahrscheinlich erscheint hier ein Zusammenhang mit der Kiefermuskulatur und der von ihr ausgehenden Kräften. Im dritten und letzten Teil dieser Reihe wollen wir anhand von einer im Rahmen meiner Disseration entstandenen Studie überprüfen, ob wir in einem fossilen Schädel Hinweise zur Entstehung von Schläfenöffnungen finden können.
Werfen wir dazu erst nochmal einen Blick auf eine untere anderem von Olson (1961) und Fox (1964) vertretene Hypothese. Demnach verteilen sich beim Beißen die Kräfte ungleichmäßig über den Schädel. In diesem „Netzwerk aus Stresslinien“ gibt es entsprechend Stellen im Schädel, in denen die Knochen kräftiger sind, um diesen Kräften standzuhalten. Andere Stellen sind entsprechend weniger kräftig oder verknöchern im Lauf der Evolution gar nicht mehr. Das Ergebnis: Schläfenöffnungen.
Fox hat hier zur Anschauung Captorhinus verwendet, eine Gattung urtümlicher Reptilien aus dem Perm der USA, deren Schädel über keinerlei Schläfenöffnungen verfügte. Laut Fox soll das Zentrum der „Wange“ dieser Tiere besonders dünn gewesen sein, was eine mögliche „Vorstufe“ für eine Öffnung darstellen könnte.
Knochennähte als Kräfteanzeiger
Diese Beobachtungen fanden auch wir spannend. In einem Paper wollten wir überprüfen, ob sich in einem Schädel ohne Schläfenöffnungen wirklich Schwachstellen nachweisen lassen und ob diese sich an den Stellen befinden, an denen andere Tiere Schläfenöffnungen entwickelt haben. Als Modellorganismus diente auch uns hierbei Captorhinus, genauer die Art Captorhinus aguti (Abb. 1). Wir haben uns für diese Tiere entschieden, da sie als frühe Amnioten relativ nahe anderen Arten mit Schläfenöffnungen standen und gleichzeitig durch eine große Zahl an dreidimensional erhaltenen Schädeln relativ gut belegt sind.

Da Öffnungen nur zwischen den Knochen entstehen, haben uns vor allem die Knochennähte interessiert. Die Schädelknochen wachsen hier nicht nur zusammen, eine Knochennaht (oder auch Sutur) dient auch dazu auf den Schädel wirkende Kräfte aufzufangen und weiterzuleiten.1 Hier muss man verstehen, dass sich die einzelnen Schädelknochen bei einem lebenden Tier nicht direkt berühren, sondern dazwischen immer eine dünne Schicht an elastischem Gewebe liegt.2
Anhand von Experimenten mit heutigen Tieren weiß man, dass die Dicke der Knochennähte sowie ihre allgemeine Gestalt in Zusammenhang mit der Art und Intensität der auf sie wirkenden Kräfte stehen.3, 4 Verallgemeinernd könnte man hierbei sagen, dass komplexere und dickere Nähte wie z. B. kräftige Verzahnungen eher auf höhere Kräfte hinweisen als einfache Überlappungen oder dünne Verbindungen entlang einer geraden Linie.
Wir können ähnliche Experimente natürlich nicht mehr mit Captorhinus durchführen. Stattdessen können wir allerdings den Weg rückwärts gehen und anhand der fossil erhaltenen Knochennähte versuchen Rückschlüsse zur einstigen Kräfteverteilung im Schädel zu ziehen.
Einen Schädel digital auseinanderbauen
Hier stoßen wir auf erste Probleme. Einmal ist das elastische Weichgewebe zwischen den Knochen fossil nicht überliefert. Wir können uns also nur auf die Struktur des knöchernen Teils der Nähte verlassen. Gleichzeitig werden wir ein unvollständiges Bild haben, wenn wir die Knochennähte nur von außen betrachten können. Klar, Verzahnungen sind oft auch auf der Außenseite sichtbar, aber wie komplex das „Innenleben“ einer Sutur ist und natürlich auch wie dick die Knochen an dieser Stelle sind lässt sich nur sehen, wenn wir den Schädel buchstäblich auseinanderbauen. Das ist besonders bei versteinerten Schädeln natürlich schwierig.
Wenn wir nicht gerade das Glück haben, dass ein Schädel vor seiner Fossilisation in seine Einzelteile zerfallen ist, helfen hier nur die Wunder der Technik. Dank unseres Ko-Autoren David Ford konnte uns hierfür der Scan eines Captorhinus Schädels zur Verfügung gestellt werden. Das Originalstück befindet sich im Sam Noble Oklahoma Museum of Natural History und wurde bereits 2017 an der University of Texas mittels Computertomografie geröntgt. Das Ergebnis ist eine lange Serie von Schnittbildern (1897 Bilder um genau zu sein) aus verschiedenen Perspektiven, die es erlauben quasi wie bei einem Daumenkino durch das Innere des Schädels zu kucken.
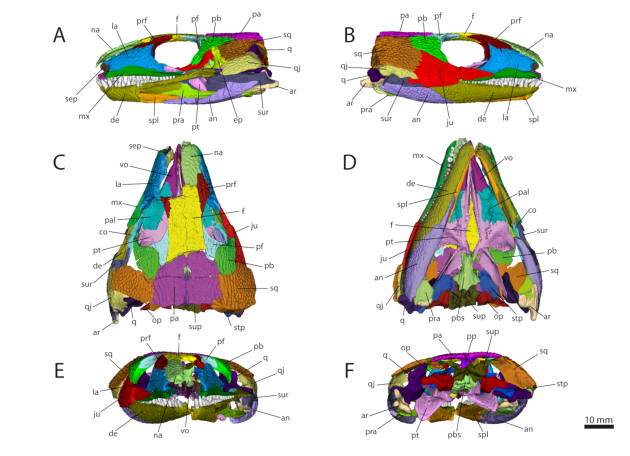
Das hilft uns schon ein großes Stück weiter. Die eigentliche Arbeit aber fängt jetzt erst an. Um aus diesen vielen Bildern ein ansehnliches 3D-Modell (Abb. 2) zu generieren, müssen nun Bild von Bild alle sichtbaren Knochen identifiziert und digital nachgezeichnet werden („Tracing“). Diese Mammutaufgabe hat aus unserem Team in erster Linie Yannick Pommery übernommen, damals Gast-Masterstudent in Tübingen. Die vielen Nachzeichnungen dann noch mit der Spezialsoftware interpolieren, fertig ist der digitale Schädel.
Mit einem passenden Programm zur Darstellung von 3D Objekten (in unserem Fall MorphoDig) können wir jetzt zum Beispiel jeden Knochen zur besseren Unterscheidung einfärben oder auch einzelne Knochen „ein- und ausschalten“, je nachdem welchen Aspekt des Schädels wir uns gerade ansehen möchten. Unser 3D Modell ist übrigens frei verfügbar auf MorphoMuseuM, wo ihr auch ohne extra Download ein bisschen damit herumspielen könnt.
Vielfalt im Captorhinus Schädel
Wie erwartet können wir nun anhand des Modells sehen, dass die Knochennähte bei Captorhinus sowohl in ihrer Gestalt als auch in ihrer Dicke variieren (Abb. 3)
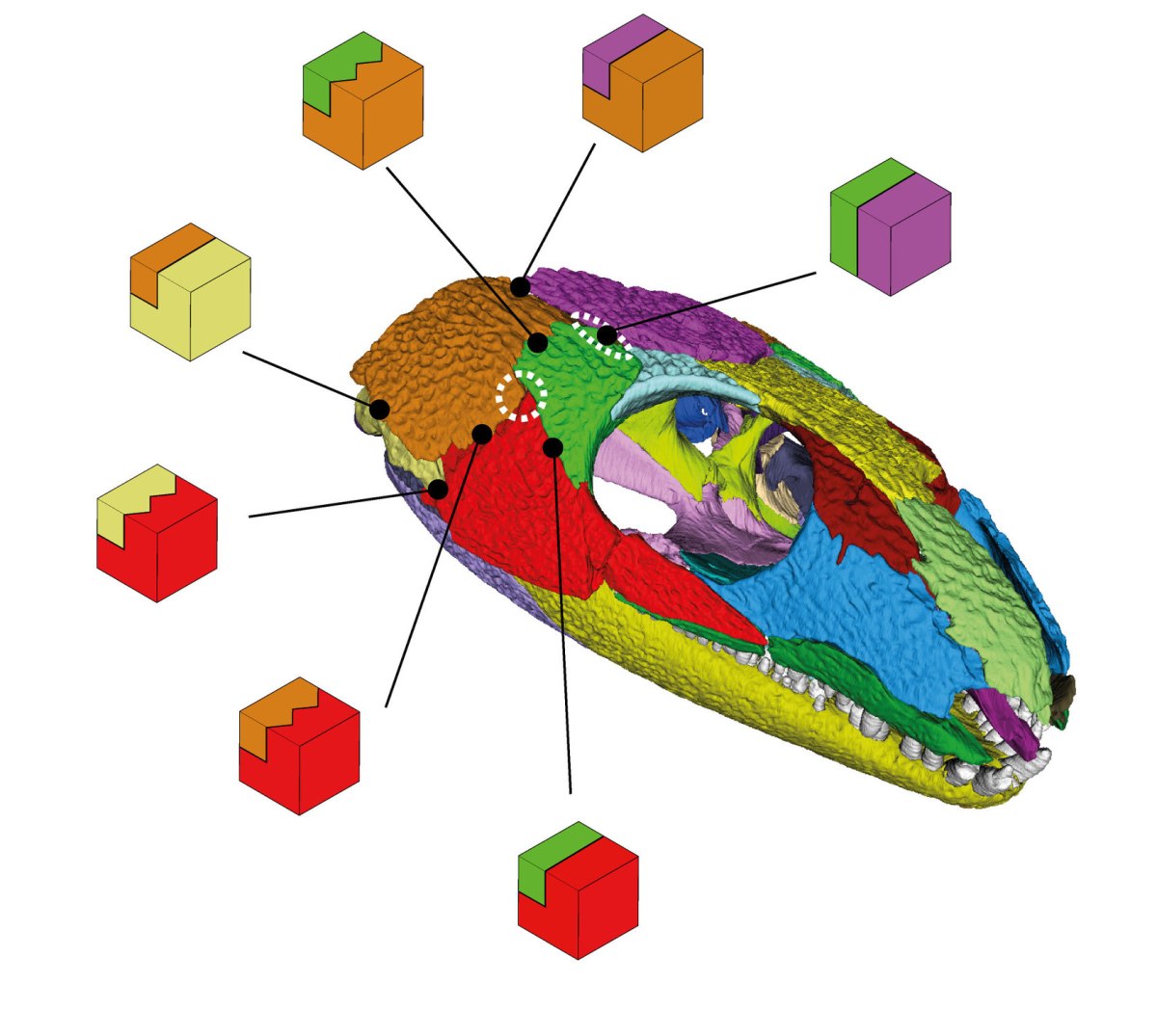
Bei den Nähten können wir insgesamt acht verschiedene Verbindungstypen unterscheiden. Die einfachste Verbindungsart ist der sogenannte „butt joint“ (Stoßfuge). Hier treffen zwei Knochen mehr oder weniger gerade aufeinander. Das sehen wir vor allem an der Längsachse des Schädels zwischen den beiden Scheitelbeinen, aber auch zwischen Scheitelbein und Postorbitale (grün-violett in Abb. 3). Letzteres wird nachher noch relevant.
Deutlich komplexer sind hier die drei Typen an Verzahnungen. Die Verzahnungen können in der Horizontalen, Vertikalen oder auch in beiden Dimensionen angeordnet sein. Letztere sehen wir insbesondere längs zwischen beiden Stirnbeinen direkt über den Augen. Hier sind auch die Knochen verhältnismäßig dick. Ein gutes Zeichen, dass diese Region größeren Kräften standhalten musste. Eventuell, weil hier sowohl die Kräfte der Kiefermuskulatur als auch der zubeißenden Schnauze gewirkt haben.
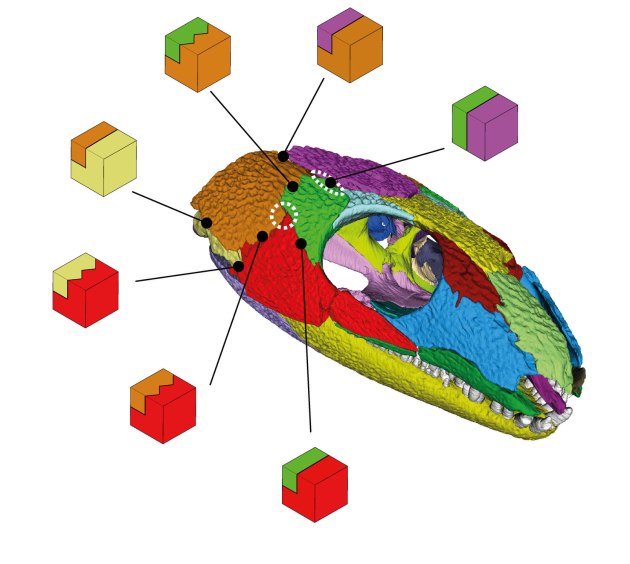
Am häufigsten im gesamten Schädel und auch vorherrschend in der Schläfenregion von Captorhinus sind allerdings die abgestuften Verbindungen. Heißt, ein Knochen bildet auf seiner Innenseite eine „Plattform“ aus, der der benachbarte Knochen aufliegt und mit seinem „Vorderende“ dann auf die äußere Hälfte der Naht des ersten Knochens trifft. Die äußere Naht kann dabei simpel wie bei einem „butt joint“ oder auch verzahnt sein.
Insgesamt fällt eine „Verschachtelung“ der Schläfenknochen auf, die die gesamte Region wohl stabiler gemacht hat. Das Schuppenbein (braun) überlappt Quadratojugale (hellgelb) und Jochbein (rot). Wird aber selbst von Scheitelbein (violett) und Postorbitale (grün) überlagert.
Interessant ist, dass die verzahnten Stufennähte, die wahrscheinlich stabiler als die einfachen Stufennähte waren, beinahe über die gesamte mittlere Vertikale durch die „Wange“ von Captorhinus verlaufen. Nur direkt im Wangenzentrum, am Treffpunkt Postorbitale, Joch- und Schuppenbein hat die Verzahnung eine Lücke (Abb. 3). Dort haben wir außerdem genau wie bereits Fox (1964) die geringste Knochendicke gemessen. Alles in allem also ein guter Hinweis, dass hier bei Captorhinus und vielleicht auch bei anderen frühen Amnioten eine Schwachstelle war (Abb. 3, gestrichelter Kreis).
Unserer Meinung nach befindet sich in der Schläfenregion von Captorhinus auch noch eine weitere Schwachstelle, nämlich in der bereits erwähnten Verbindung zwischen Scheitelbein und Postorbitale (Abb. 3, obere gestrichelte Ellipse). Die Naht ist hier zwar nicht ungewöhnlich dünn, bildet aber nur einen simplen „butt joint“, während alle anderen angrenzenden Nähte komplexer sind.
Rekonstruktion der Kiefermuskulatur
So weit so gut. Bestimmung möglicher Schwachstellen ist das eine. Würde das aber auch mit der tatsächlichen Bewegung der Kiefermuskulatur zusammenpassen? Wie zuvor erwähnt, interessieren uns hier besonders Kieferadduktoren, also jene Muskeln, die den Kiefer schließen.
Auch hier haben wir das Problem, dass unsere Kenntnisse zwangsläufig begrenzt sind. Muskulatur bleibt fossil in der Regel nicht erhalten. Wir müssen uns also wieder rantasten. Ein guter Hinweis für die Anordnung von Muskeln sind das Vorhandsein verschiedener Fortsätze, Kämme oder Narben als Ansatzstellen an den Knochen.5 Allerdings können Muskeln auch an Knorpeln oder anderem Weichgewebe ansetzen, das wie Muskeln nur selten erhalten bleibt.6
Ein zweiter Erkenntnisweg könnte hier über die nächsten lebenden Verwandten sein. Bei Captorhinus sind das aufgrund seiner basalen Stellung die Gesamtheit der heutigen Reptilien einschließlich der Vögel (Abb. 4). Problem nur, heutige Reptilien sind entgegen ihrem Ruf nicht „urtümlich“ und haben je nach Gruppe ihre Schädelanatomie deutlich verändert. Man müsste also über die Gemeinsamkeiten der verschiedenen Gruppen versuchen den Urzustand ihres letzten gemeinsamen Vorfahren zu rekonstruieren.
Ein weiteres Problem ist die Tatsache, dass wir heute schlicht keine Tiere mehr haben, die den ursprünglichen Schädel ohne Schläfenöffnungen beibehalten haben. Wir können also erstmal nicht wissen, wie sich die Kieferadduktoren in so einem Schädel anordnen würden. Abhilfe könnte hier zum Beispiel der Blick auf die Muskulatur heutiger Meeresschildkröten schaffen, da diese ihre Schläfenöffnungen wieder zurückgebildet haben.7
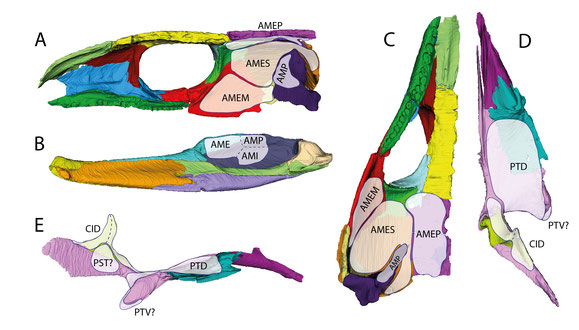
Kombinieren wir den vermuteten Urzustand der heutigen Reptilien8 mit unseren Kenntnissen zur Anordnung der Kiefermuskulatur bei Meeresschildkröten und den anatomischen Details des Captorhinus Schädels ergibt sich folgendes Modell (Abb. 5):
Captorhinus aguti hatte einen dreigeteilten äußeren Kieferadduktor (AME), der große Flächen der inneren Wange (AMEM, AMES) und des inneren Scheitelbeins (AMEP) einnahm. Der innere Kieferadduktor (AMI) nahm vor allem die Oberseite des knöchernen Gaumens (PTD) ein. Eventuell war auch ein unterer Teil (PTV) mit dem Pterygoid („Flügelbein“) verbunden. Der hintere Adduktor (AMP) setze am Quadratum an. Am Gaumen haben außerdem ein constrictor internus dorsalis und eventuell ein pseudotemporalis angesetzt.
Es sollte auch erwähnt werden, dass wir nicht die ersten sind, die die Kiefermuskulatur von Captorhinus und anderen Captorhiniden rekonstruiert haben. Wir hatten mit unserer Studie insbesondere die Modelle von Fox (1964) und Heaton (1979) erweitert.
Die Entstehung von Schläfenöffnungen
Passt unsere Rekonstruktion der Kieferadduktoren mit der Gestalt der Knochennähte zusammen? Ich meine ja. Im Grunde würden wir in der Schläfenregion von Captorhinus während dem Zubeißen zwei Hauptbewegungen sehen für die jeweils die äußeren Kieferadduktoren (AME) verantwortlich sind (Abb. 6).
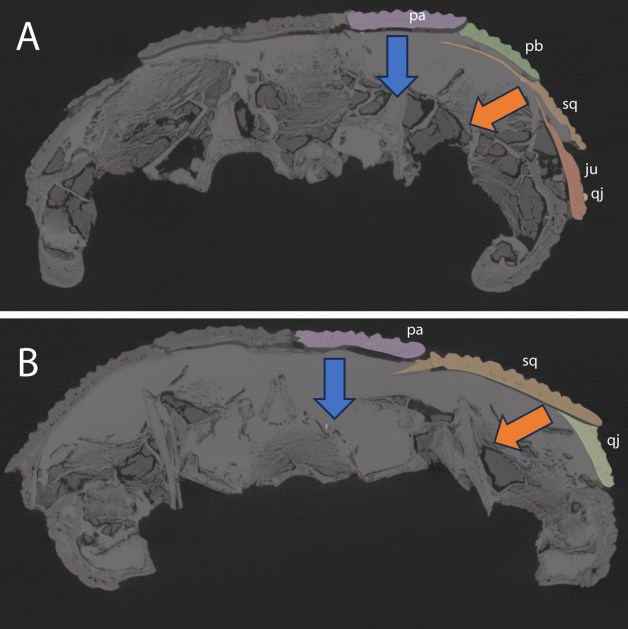
Einmal haben wir die oberflächlichen und mittleren Teile (AMES und AMEM), die beim Schließen des Kiefers die „Wangenknochen“ (Jochbein, Schuppenbein, Postorbitale und Quadratojugale) nach innen ziehen. Das könnte erklären, warum diese untereinander „verschachtelt“ sind (siehe oben). Eine solche Anordnung dürfte die Knochen bei diesem Zug zusammengehalten haben.
Intuitiv würde ich erwarten, dass hier der schwächste Punkt tatsächlich etwa in der Mitte der Wange liegen würde. Dafür sprechen auch Knochenleisten an den inneren Rändern der Wange, an die wohl der kräftige sehnige Teil der Kieferadduktoren angesetzt hat (auch bereits von Fox 1964 berichtet).
Das Herunterziehen des Scheitelbeins durch den AMEP wurde wohl durch die Stufennaht mit dem Schuppenbein stabilisiert. Auch die Verzahnungen in Richtung Stirnbein und Postfrontale dürften zur Stabilisierung beigetragen haben. Kann sein, dass sich die mutmaßlich schwache Verbindung zwischen Scheitelbein und Postorbitale durch die mittige Position zwischen den beiden „Stabilisatoren“ erklären lässt und hier kaum Muskeln ansetzten. Ich würde aber nicht behaupten, hier eine eindeutige Erklärung zu haben.
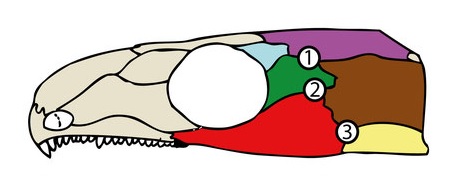
Grundsätzlich passen die beiden „Schwachstellen“ im Captorhinus Schädel zum Teil mit der Position der Schläfenöffnungen anderer früher Amnioten überein (Abb. 7). Unsere Schwachstelle zwischen Postorbitale und Scheitelbein (1 in Abb. 7) entspricht weitesgehend dem oberen Schläfenfenster vieler Diapsiden. Der Hauptunterschied liegt darin, dass in der Regel auch das Schuppenbein und je nach Art einige weitere Knochen den Rand des Fensters bilden.9
Ähnlich verhält es sich mit der Schwachstelle in der Wange (2 in Abb. 7). Einige frühe Synapsiden wie Dimetrodon (Abb. 2C im ersten Teil) hatten wie hier eine Öffnung exakt zwischen Jochbein, Schuppenbein und Postorbitale. Das dürfte allerdings auch daran liegen, dass diese Arten ihr Quadratojugale bereits deutlich zurückgebildet haben. Letzteres ist bei vielen anderen frühen Amnioten ebenfalls Teil der Umrandung. Manchmal erstreckt sich die Öffnung sogar bis zum Scheitelbein hoch.9
Lediglich für eine Öffnung zwischen Jochbein, Schuppenbein und Quadratojugale (3 in Abb. 7) finden sich keine Anhaltspunkte bei Captorhinus. Die kommt vor allem bei den ausgestorbenen Parareptilien vor.10
Alles wirklich so einfach?
Ich denke nicht, dass die Einbeziehung weiterer Knochen zur Umrandung der Schläfenöffnungen der „Schwachstellen-Hypothese“ widerspricht. Vielmehr sollte man diese Stellen im Schädel als ersten Ausgangspunkt für eine Öffnung sehen, wie bereits in Teil 2 zu dem frühen Synapsiden Ophiacodon diskutiert. Weitere Spezialisierungen und etwaige Vergrößerungen der Kieferadduktoren könnten die Schläfenöffnungen im Anschluss auf andere Knochen erweitert haben. Alternativ könnten größere Öffnungen auch durch eine Verschmelzung zweier Schwachstellen entstanden sein (siehe wieder Ophiacodon).
Auch das Fehlen einer Schwachstelle bei „Position 3“ im Captorhinus Schädel muss nicht viel heißen. Die Anordnung der Kieferadduktoren variiert je nach Art, entsprechend können auch die Schwachstellen leicht woanders liegen. Es kann gut sein, dass ein Vorfahre der erwähnten Parareptilien dadurch an „Position 3“ seinen Schwachpunkt hatte.

Ein Problem könnte hingegen Captorhinus selbst sein. Zwar gibt es gute Gründe, warum wir das Tier als Modellorganismus verwendet haben, es muss aber erwähnt werden, dass Captorhinus aguti bereits etwa 30 Millionen Jahre nach den ältesten bekannten Amnioten gelebt hat.10 Selbst Arten mit Schläfenöffnungen gab es schon deutlich vor dem Erstauftreten von Captorhinus.
Erwartbar hatten bei Captorhinus seitdem Veränderungen stattgefunden. Das betrifft vor allem die Spezialisierung des Kiefers und der Zähne zu einer allesfressenden, wenn nicht sogar rein pflanzenfressenden Lebensweise11 (im Gegensatz zu den wahrscheinlich insektenfressenden „Ur-Amnioten“)12. „Fortschrittlichere“ Captorhiniden wie Labidosaurus (Abb. 8) oder die Moradisaurinen hatten diese Spezialisierungen weiter geführt und auch ihre Kiefermuskulatur deutlich vergrößert.13 Doch bis heute kennen wir keinen Captorhiniden, der Schläfenöffnungen entwickelt hat.
Die Sache bleibt also kompliziert. Es gibt verschiedene Möglichkeiten von hier weiter zu machen. Um den Interpretationen zur Kräfteverteilung im Schädel mehr Substanz zu geben, würde sich empfehlen für Captorhinus eine Finite-Elemente-Analyse durchzuführen wie es bereits für viele andere Gruppen gemacht wird.14 Auch sollte man definitv einen Blick auf andere frühe Amnioten werfen, vor allem jene, die noch „urtümlicher“ als Captorhinus und gleichzeitig nahe Verwandte von Arten mit Schläfenöffnungen waren (z. B. Protorothyris). Zuletzt müsste man sich mehr Schädel von Captorhinus aguti ansehen, um zu zeigen, dass die hier vorgestellten Muster bei der Art auch wirklich die Regel sind.
Es bleibt spannend.
Weiterführende Literatur
- Abel, P., Pommery, Y., Ford, D.P., Koyabu, D. und Werneburg, I. (2022). Skull sutures and cranial mechanics in the Permian reptile Captorhinus aguti and the evolution of the temporal region in early amniotes. Frontiers in Ecology and Evolution 10: 841784. doi: 10.3389/fevo.2022.841784
- Werneburg, I. und Abel, P. (2022). Modelling skull network integrity at the dawn of amniote diversification with considerations on functional morphology and fossil jaw muscle reconstructions. Frontiers in Ecology and Evolution 9: 799637. doi: 10.3389/fevo.2021.799637
Andere erwähnte Referenzen
- Fox, R. C. (1964). The adductor muscles of the jaw in some primitive reptiles. University of Kansas Publications 12, 657–680. https://www.gutenberg.org/files/30321/30321-h/30321-h.htm
- Heaton, M. J. (1979). Cranial anatomy of primitive captorhinid reptiles from the Late Pennsylvanian and Early Permian Oklahoma and Texas. Oklahoma Geological Survey 127, 1–84. doi: 10.1130/spe152-p1
- Olson, E. C. (1961). Jaw mechanisms: rhipidistians, amphibians, reptiles. American Zoologist 1, 205–215. doi: 10.1093/icb/1.2.205

{kind=link}