

Titelbild: Skelett einer Brückenechse (Sphenodon punctatus) in der Paläontologischen Sammlung Tübingen. Deutlich zu sehen die zwei Paar Schläfenöffnungen. Foto: Agnes Fatz
Habt ihr euch die Schädel von Wirbeltieren einmal genauer angesehen? So ein Schädel hat verschiedene Öffnungen, am prominesten davon in der Regel die Augenhöhlen und die Nasenlöcher. Besonders bei Reptilien wie der Brückenechse hier im Titelbild befinden sich oft noch weitere Öffnungen im Bereich der Schläfe. Bei der Brückenechse sind es zwei Paare, eins an der Seite und ein zweites eher in Scheitelnähe. Diese Konstellation mit zwei Paar Schläfenöffnungen nennt man auch „diapsid“ („zwei Knochenbögen“). Die Tiergruppe, bei der dieser Schädeltyp am häufigsten vorkommt heißt entsprechend Diapsida.
Die Diapsida umfasst im Grunde alle noch lebenden Reptilien, inklusive der Vögel. Der typische „diapside“ Schädel kommt heute neben der Brückenechse auch bei den meisten Krokodilen vor (Abb. 1A,C). Die anderen Diapsiden haben diesen Typ allerdings mehr oder weniger abgewandelt (Abb. 1B).
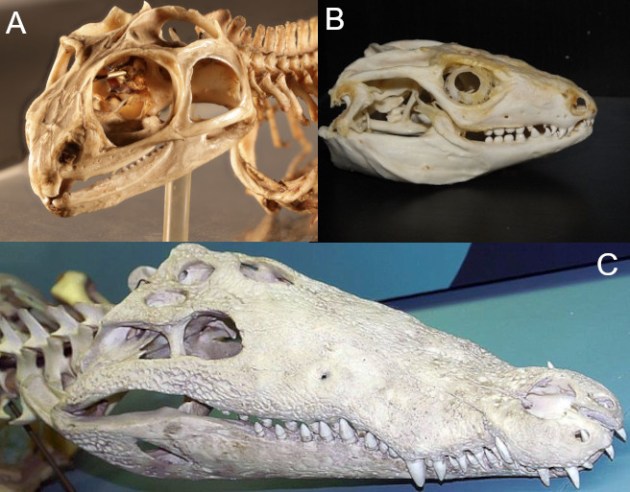
Bilder: A von Agnes Fatz; B von HCA (CC BY-SA 4.0); C von David J. Stang (CC BY-SA 4.0).
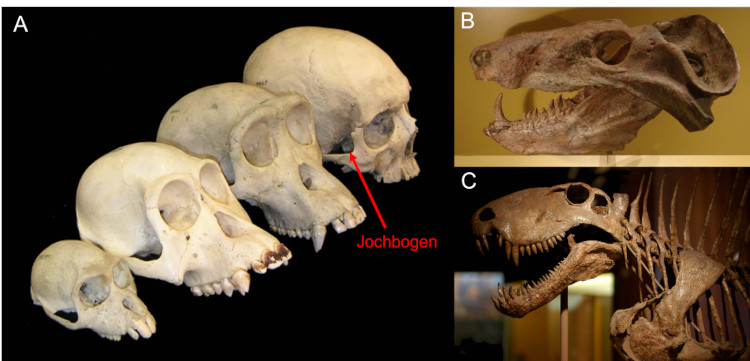
Auch wir Menschen haben Schläfenöffnungen (Abb. 2A). Die sind bei uns etwas anders positioniert, anatomisch Versierte kennen ihre Umrandung als Jochbogen. Dass diese mit den Schläfenöffnungen der Reptilien vergleichbar sind, zeigt sich wenn wir die Zeit etwas zurückdrehen. Ausgestorbene Verwandte der Säugetiere wie die Cynognathier der Trias (Abb. 2B) hatten auch einen deutlich ausgeprägten Jochbogen, aber man erkennt auch gut das Paar großer Schläfenöffnungen. Gehen wir noch weiter zurück in die Perioden Perm und Karbon, zu den frühesten Vorfahren der Säugetiere, erkennt man, dass sich die Schläfenöffnungen der Säugetiere über die Zeit horizontal verbreitert hatten. Ursprünglich hatten auch unsere Vorfahren ähnlich den Reptilien ein seitliches Paar Schläfenöffnungen, direkt hinter den Augen (Abb. 2C)
Diesen Schädeltyp, bei dem nur die seitlichen Schläfenöffnungen vorhanden sind nennt man „synapsid“ („mit einem Knochenbogen“). Die gemeinsame Gruppe der Säugetiere samt all ihrer ausgestorbenen Verwandten heißt passend Synapsida.
Doch warum überhaupt haben viele Wirbeltiere diese Öffnungen? Warum sind sie oft unterschiedlich ausgeprägt? Und ist es wirklich sinnvoll Wirbeltiere anhand der Anzahl ihrer Schläfenöffnungen zu kategorisieren? Besonders um die letzte Frage soll es in Teil 1 dieser dreiteiligen Artikelreihe gehen.
Eine lange Forschungsgeschichte
Der Evolution dieser Schläfenöffnungen hatte ich mich ausgiebig in meiner Dissertation gewidmet.1,2,3 Dabei reihe ich mich in eine lange Liste von WissenschaftlerInnen, die sich ebenfalls mit den oben genannten Fragen beschäftigt haben. Erste Arbeiten, die sich detailliert mit anatomischen Unterschieden innerhalb der Schläfenregion auseinandergesetzt haben, gab es dabei bereits vor über 180 Jahren.4 Einen frühen Peak erlebte das Thema am Übergang in das 20. Jahrhundert als die Evolutionstheorie Darwins langsam Fuß fasste.5,6,7,8
In diese Zeit fiel auch die Benennung der Gruppen Synapsida und Diapsida durch Henry Osborn im Jahr 1903.9 Dieser schlug dadurch eine Zweiteilung der „Reptilien“ vor. Seine Synapsiden und Diapsiden hatten dabei noch eine etwas andere Zusammensetzung. Anders als heute waren für ihn zum Beispiel auch Schildkröten und Plesiosaurier Synapsiden, also Verwandte der Säugetiere.

Schädel nach Fox & Bowman (1966). Silhouetten (von oben links im Uhrzeigersinn) von Dmitry Bogdanov/Roberto Díaz Sibaja (CC BY 3.0), Christoph Schomburg, Gareth Monger (CC BY 3.0), Frank Denota, Steven Traver, Nobu Tamura/T. Michael Keesey (CC BY-SA 3.0), Dmitry Bogdanov (CC BY 3.0), Brad McFeeters/T. Michael Keesey, T. Michael Keesey und Nobu Tamura/A. Verrière (CC BY-NC-SA 3.0).
Dieses Schema wurde von anderen WissenschaftlerInnen aufgenommen und erweitert (Abb. 3). Eine der bekanntesten Veränderungen kam von Samuel Williston in 1917, der zwei weitere Gruppen ergänzte: die Anapsida („ohne Knochenbögen“) und Parapsida (etwa „nebenstehend mit Knochenbögen“). 10 Die Anapsiden waren nach Williston die Stammgruppe der meisten anderen „Reptilien“ und besaßen selbst noch keine Schläfenfenster. Die Schildkröten wären als eine Art Relikt die einzigen heutigen Vertreter dieser Gruppe. Die Parapsiden wiederum könnte man als dritte Gruppe von „Reptilien“ mit Schläfenfenstern verstehen, die sich nach Williston unabhängig der Synapsiden und Diapsiden entwickelt hätten. Williston zählte dazu unter anderem die ausgestorbenen Mesosaurier und Ichthyosaurier, sowie die Schuppenechsen (Eidechsen, Schlangen etc.).
Ebenfalls erwähnenswert ist der Begriff Euryapsida („breite Knochenbögen“), der 1945 von Edwin H. Colbert für eine Gruppe aus unter anderem Plesiosauriern und Pflasterzahnsauriern eingeführt wurde.11
Neben diesen taxonomischen Bezeichnungen, welche auf der Annahme basierten, dass Arten mit einer scheinbar gleichen Gestalt der Schläfenregion nahe miteinander verwandt waren, wurden im Lauf der Jahrzehnte viele weitere eher beschreibende Begriffe eingeführt. Darunter zum Beispiel stegokrotaph („bedeckte Schläfe“) und gymnokrotaph („nackte Schläfe“),6 die heute noch Anwendung bei der vergleichenden Anatomie der Amphibien finden.12
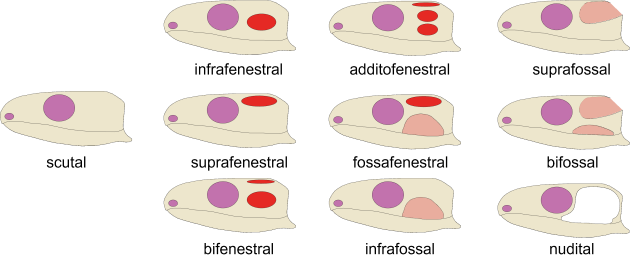
Seit dem Ende des 19. Jahrhunderts wurden etwa 20 verschiedene Systeme zur Einteilung der verschiedenen Schädeltypen eingeführt. Die hier alle vorzustellen, würde gewaltig den Rahmen sprengen. Wer hier tiefer eintauchen möchte, dem empfehle ich unsere Arbeit in Biological Reviews. Dort stellen wir ebenfalls ein komplett neues System vor, nach dem sich unter allen Landwirbeltieren zehn Haupttypen in der Schläfengestalt idenfizieren lassen (Abb. 4).
Wie aussagekräftig ist die Anzahl der Schläfenöffnungen?
Bei all den Einteilungsmöglichkeiten und aufgestellten Namen wie Synapsida, Anapsida usw. stellt sich natürlich die Frage, ob es überhaupt sinnvoll ist, die großen Gruppen der Landwirbeltiere anhand ihrer Schläfengestalt zu identifizieren. Zweifel daran gibt es schon lange.13,14 Durch viele neue Fossilfunde sowie durch die Etablierung umfangreicher computergestützter Verwandtschaftsanaylsen ist mittlerweile klar: Die Sache ist bisschen komplizierter (Abb. 5).
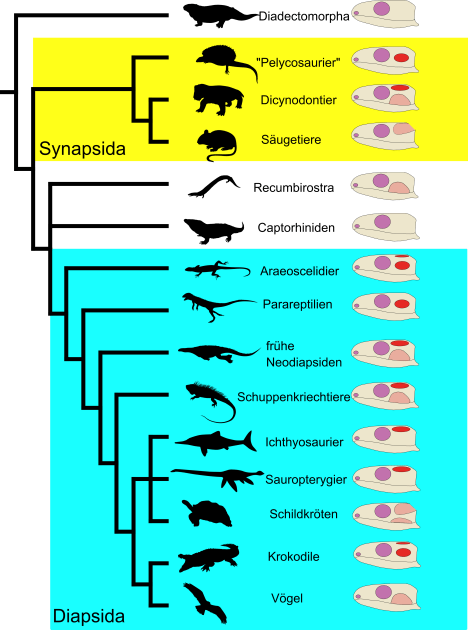
Silhouetten (v.o.n.u.) von Dmitry Bogdanov/Roberto Díaz Sibaja (CC BY 3.0), Dmitry Bogdanov (CC BY 3.0), Matt Celeskey, Soledad Miranda-Rottmann (CC BY 3.0), Smokeybjb (verändert, CC BY-SA 3.0), Dmitry Bogdanov (verändert, CC BY-SA 3.0), Nobu Tamura/T. Michael Keesey (CC BY-SA 3.0), Nobu Tamura (CC BY-NC-SA 3.0), Nobu Tamura/T. Michael Keesey (CC BY-SA 3.0), Jack Mayer Wood, Gareth Monger (CC BY 3.0), Arthur S. Brum, Cathy (CC BY-NC-SA 3.0), Steven Traver, Christos Barboutis.
Die meisten WissenschaftlerInnen sind sich weiterhin einig, dass es eine große Säugetiergruppe (Synapsida) und eine Gruppe um die heutigen Reptilien und Vögel (Diapsida) gibt. Alle anderen wie die Anapsida, Parapsida und Euryapsida dagegen scheinen keine natürlichen Gruppierungen zu sein.
Ein prominentes Beispiel hier sind Schildkröten. Statt ein heutiges Relikt „anapsider“ Reptilien zu sein, sprechen die meisten anatomischen und genetischen Analysen dafür, dass Schildkröten in Wahrheit Diapsiden sind. Konkret, der „anapside“ (oder nach unserem Schema) „scutale“ Schädel der Schildkröten entwickelte sich aller Wahrscheinlichkeit nach aus einem diapsiden Vorfahren mit Schläfenfenstern (Abb. 6).15,16
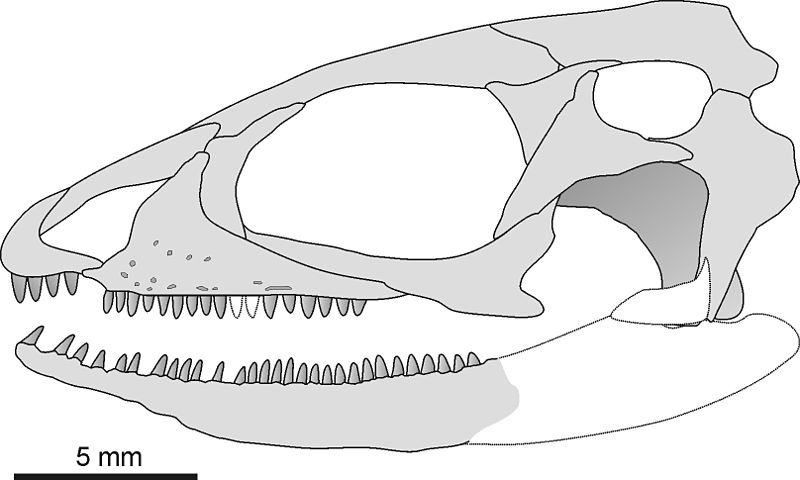
Auch der klassische „synapside“ (=infrafenestrale) Schädel kommt neben den namensgebenden Synapsiden unter anderem ebenfalls bei den ausgestorbenen Parareptilien, sowie heutigen Glattstirnkaimanen und einigen Fröschen vor.17,18, 19
Gleichzeitig wissen wir, dass sich der Schläfentyp auch innerhalb einer Art ändern kann. Bei den eben erwähnten Parareptilien z.B. kennen wir einige Fälle, bei denen sich ein bei den Jungtieren vorhandenes Schläfenfenster im Laufe der Zeit schließt oder zweigeteilt wird.20,21 Bei anderen Arten, darunter auch frühe Synapsiden, tauchen gelegentlich zusätzliche Schläfenfenster auf, manchmal auch nur an einer Seite des Schädels.22,23
Alles in allem dürfte klar sein, dass die Evolution der Schläfenfenster komplex ist und eine Einteilung der Landwirbeltiere anhand ihrer Schläfengestalt nicht unbedingt sinnvoll ist. Aber warum sehen wir diese komplexe Evolution und warum entstanden manche Typen mehrmals unabhängig voneinander?
Mehr dazu im zweiten Teil.
Weiterführende Literatur
- Abel, P. und Werneburg, I. (2021). Morphology of the temporal skull region in tetrapods: research history, functional explanations, and a new comprehensive classification scheme. Biological Reviews 96 (5): 2229–2257. doi: 10.1111/brv.12751
- Abel, P. und Werneburg, I. (2024). The temporal region of the tetrapod skull: a textbook example on integrative morphology. Revue de Paléobiologie. https://www.researchgate.net/publication/378971484_The_temporal_region_of_the_tetrapod_skull_a_textbook_example_on_integrative_morphology
- Ford, D. P. (2018). The evolution and phylogeny of early amniotes. Dissertation, University of Oxford.
- Ford, D. P. und Benson, R. B. J. (2020). The phylogeny of early amniotes and the affinities of Parareptilia and Varanopidae. Nature Ecology & Evolution 4(1), 57–65. doi: 10.1038/s41559-019-1047-3
- Werneburg, I. (2013). The tendinous framework in the temporal skull region of turtles and considerations about its morphological implications in amniotes: a review. Zoological Science 30(3), 141–153. doi: 10.2108/zsj.30.141
- Werneburg, I. (2019). Morphofunctional categories and ontogenetic origin of temporal skull openings in amniotes. Frontiers in Earth Science 7, 1–7. doi: 10.3389/feart.2019.00013
Andere erwähnte Referenzen
- Colbert, E. H. (1945). The Dinosaur Book: The Ruling Reptiles and their Relatives. The American Museum of Natural History, New York. doi: 10.5962/bhl.title.68178
- Fox, R. C. & Bowman, M. C. (1966). Osteology and relationships of Captorhinus aguti (Cope) (Reptilia: Captorhinomorpha). The University of Kansas Paleontological Contributions Vertebrata 11, 1–79. https://www.semanticscholar.org/paper/Osteology-and-relationships-of-Captorhinus-aguti-Fox-Bowman/0fe77983215c9c01c52a77b94ad6f8460c978135
- Osborn, H. F. (1903). On the primary division of the Reptilia into two sub-classes, Synapsida and Diapsida. Science 17(424), 275–276. doi: 10.1126/science.17.424.275.
- Williston, S. W. (1917). The phylogeny and classification of reptiles. The Journal of Geology 25(5), 411–421. doi: 10.1086/622507

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}